
This week we’re finishing up our in-depth look at energy metabolism. In the first segment of this series, we looked closely at the biochemistry of energy metabolism and focused specifically on the equations of the phosphagen system, glycolytic system, and aerobic carbohydrate and fat metabolism. In the second segment, we looked at what does and does not cause muscular acidosis. Now, in this segment, we’re going to look at how fats and carbs differ as fuels and how that influences their utility.
In my opinion, this is one of the biggest areas where a deep understanding (or at least a deeper understanding) can dispel pseudoscientific sports science before it really begins. When we start talking about fats and carbohydrates and how they’re used within the context of exercise, we’re really talking about fuel capabilities—and these two fuels have radically different biochemical profiles! It’s these profiles that lead to their relative advantages and disadvantages, and whenever someone claims that this or that fuel can overcome those built-in biochemical differences we know we’re dealing with a charlatan.
The only other thing I’m going to say to preface this final segment is to keep in mind the context. Right here, we’re talking about the basic attributes of fuels based on their biochemistry. In the wider context of human physiology and adaptability, we might see athletes compensate (not “overcome”) for these limitations. I’ll reiterate this further throughout, but for now let’s move onto the points themselves.
#1: Fat Is Better for Storage but Worse for Energy Regeneration
Fat is an excellent storage molecule. Not only is it much denser in energy than carbohydrates (9 kcal per gram for fat vs. 4 kcal per gram for carbs), it also doesn’t require water to be stored. Thus, it makes perfect sense why our body stores energy surpluses as fat and not as carbohydrate. It’s not because fat is the “perfect energy source”—it’s because fat is the perfect storage molecule!
When it comes to regenerating ATP in a performance context, however, fat loses against carbohydrates on two counts:
Fats Are Slow
Metabolizing a molecule of fat requires more reactions than aerobically metabolizing a carbohydrate (which itself requires more steps than anaerobic carb metabolism, which requires more steps than creatine phosphate metabolism). The complexity of fat metabolism ensures that it is always the slowest method of regenerating ATP—which is fine when we’re talking about fueling our day-to-day life, but problematic when we need to maximize ATP regeneration during exercise.
We’re always burning a mix of fuels when we exercise, and we can only store limited amounts of carbohydrates anyway, so in practical terms we’re not talking about a two-fold difference between metabolizing carbs versus fats—it’ll be far smaller. The more available carbs are, however, the more energy you’ll be able to regenerate moment-to-moment, and that impacts performance.
Fats Are Oxygen-Inefficient
Unlike carbohydrates, fats do not contain any oxygen atoms in the fuel-part of their molecule. That means aerobically metabolizing a molecule of fat requires more oxygen than aerobically metabolizing a molecule of carbohydrates—about 10% more oxygen.
Since climbing so frequently occludes muscles due to its isometric nature, oxygen is actually less available (to the muscles) than would be assumed based on breathing and heart rate. When muscles regain blood flow, they have a limited amount of oxygen to work with before they become occluded once again, and thus oxygen efficiency is greatly important.
This isn’t to say fats are not an important fuel source or that climbers cannot perform well without carbohydrates, as neither are true. Rather, what this suggests is that carbohydrates have inherent performance advantages over fats that are built into the molecules themselves and their metabolism. If you choose to limit or eliminate carbs from your diet, you lose these advantages without gaining any (performance advantages) in return, which isn’t a recipe for climbing success.
Aerobic Metabolism Is Always Occurring
The interplay of energy systems is often illustrated as linear:
- You begin with simple ATP hydrolysis, which lasts for a couple seconds.
- You then shift to creatine phosphate, which lasts another 7-12 seconds.
- After creatine is exhausted, you shift to anaerobic glycolysis, which lasts another 45-90 seconds.
- Finally, you reach aerobic oxidation, which will carry you indefinitely (or until you succumb to fatigue).
Simple, but Inaccurate

This is a convenient simplification, but it misses the big picture that all these energy systems are constantly working together to regenerate ATP. There is no “passing of the baton” from one system to another, there is only the increasing reliance on faster and more powerful (but more limited) systems.
Your body is fairly good at monitoring how much energy it is using and recruiting the proper systems to keep ATP at the correct level. When energy needs are low, there’s no need to recruit fast and limited systems and so the aerobic system dominates (particularly aerobic fat metabolism). When energy needs are extreme, aerobic metabolism is still occurring, but the amount of ATP it regenerates is dwarfed by the amount of ATP regenerated by the faster anaerobic systems.
In this way, time is actually the wrong measure for which system is used when; the proper measure is exertion. Since your level of exertion predicts how long you can exert yourself for, there is some correlation—but presenting time as the primary function by which your body determines energy system use is inaccurate. Let me illustrate:
Right now, as I’m typing this article out, I have very low energy needs, and those needs are easily met by aerobic fat oxidation; for every ATP hydrolyzed, there is an abundance of oxygen to metabolize a molecule of fat through the citric acid cycle to regenerate it. If I close my laptop and start walking, those needs will still be easily met, but a greater percentage of my aerobic capacity will be used, and carbs may be burned at a greater rate (though this will heavily depend on diet and so is variable person to person).
As I increase my walking speed to a jog and then a run, I will eventually maximize use of my aerobic systems (determined by my VO2 max, cardiac strength, and mitochondrial density) and begin to draw lightly upon the anaerobic systems, primarily anaerobic glycolysis. At this point, I now have a “timer” on how long I can exercise at the current intensity. If I’m only slightly above my anaerobic threshold, it may be an hour or two, but eventually my anaerobic sources will be exhausted and I will literally not have the ability to regenerate enough ATP to maintain my current level of exertion.
Picking up the pace, my ATP needs grow greater and greater beyond the capacity of the aerobic system. I still have the exact same amount of oxygen coming in (because my respiration rate cannot increase further), but the amount of ATP I can regenerate by running carbs and fat through the citric acid cycle isn’t enough to maintain my high level of exertion and so the remainder is provided by anaerobic systems. As I near an all-out sprint, my energy needs are now so high that aerobic fuel oxidation is only able to supply a small percentage of my total fuel needs.
Thus, the true picture for how the energy systems play together might look something more like this:
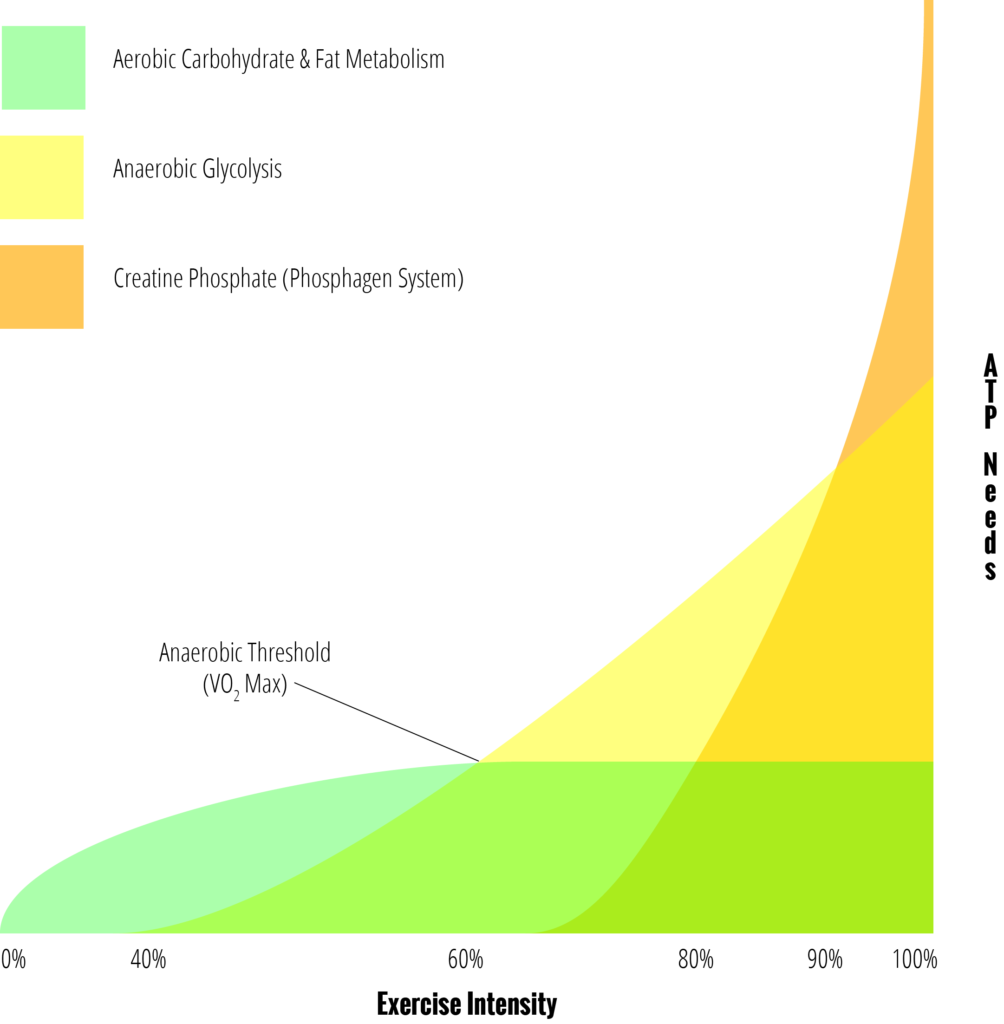
There’s never a moment when we stop using our aerobic system—or even really when we begin to use our anaerobic glycolytic or phosphagen systems—there’s only moments when the different systems provide more or less of our total ATP regeneration. At rest or a low exercise intensity, the aerobic system is more than enough to meet our needs and any ATP regenerated by creatine phosphate or anaerobic glycolysis is minimal (or, perhaps more accurately, they’re still being used but the anaerobic portion of glycolysis will always be finished with the aerobic portion and all creatine phosphate molecules that are hydrolyzed are near-instantly regenerated by the aerobic system). At high intensities, we are generating the same amount of ATP from our aerobic system but it’s not enough to meet our needs and so other systems are being used as well. The transitions are smooth, not jagged.
Shrinking One System Doesn’t Increase Another
One final insight and then this series is finished: when you shrink or eliminate one system, you do not get much compensation (if any).
Since our systems work together commensurate with our current level of exertion (see “Aerobic Metabolism Is Always Occurring” above), and since they’re limited by their capabilities (see “Fat Is Better for Storage but Worse for Energy Regeneration”, also above), we cannot increase any system by decreasing another—there’s no scale shifting.
Solely within the aerobic system, we can shift our preferred fuel from carbs to fat (or the other way around), but that doesn’t change the total amount of energy we can produce using the aerobic system so it’s a zero-sum game. Picture it like this:

As we shift the ratio of fats to carbohydrates in our diet, our body shifts from preferring one fuel to the other—but never does the total size of that energy pool change. We merely train our body to burn one fuel better while it simultaneously “forgets” how to burn the other fuel.
In a similar way, if we reduce the capacity of any system, our other systems do not “make up” for that loss. Yes, they will compensate to the extent that they can, but your total effective energy pool size will only be reduced. If you lost a lung, your VO2 max would shrink and your aerobic energy pool would shrink with it. At some levels of exertion, you’ll still be able to fully meet your energy needs, but as your energy needs increase you will eventually hit a wall. A similar (if much less devastating) thing would occur if you cut carbohydrates out of your diet—you would be able manage fine below a certain level of exertion, but the reduced total energy pool size will start to trip you up when your exercise intensity demands more ATP regeneration.
The reciprocal is also true, though: you can increase certain energy pools without decreasing others. If you train and increase your aerobic capacity, your anaerobic potential remains the same, but since you can now regenerate more ATP using the aerobic system you rely on the other systems less at a given level of intensity. Similarly, if you supplement with creatine, you increase your pool of creatine phosphate, allowing for slightly greater amounts of high-intensity exercise—you still only use that system when the situation calls for it, and your aerobic capability remains the same.
Taken together, this is probably the biggest reason why I cannot support ketogenic diets for performance—you reduce the size of one energy pool (anaerobic glycolysis) while merely shifting fuel use around in another. There is no advantage to ATP regeneration because the ketone bodies ketosis creates must also be aerobically oxidized. Thus, it’s difficult to imagine a way in which ketosis would improve performance, and as of yet no study has demonstrated any advantage to a ketogenic diet while a few have noted significant decreases in anaerobic ability—which is exactly what we would expect!
Conclusion
That wraps up this series on energy metabolism. It’s a fascinating topic, and I’m sure I’ll write more about it in the future, drawing even more insights out of this base knowledge! For now, it’s time to move on.
From this final installment, I hope you’re walking away with the following key points:
- Fats and carbs have different molecular structures and their fueling advantages and disadvantages are derived from these differences.
- Level of exertion is the most important factor in fuel selection.
- You cannot increase your aerobic system through diet, but you can affect the sizes of your anaerobic systems.
As always, I’d love to hear your thoughts! If there’s anything that is still a little unclear (or that seems contradictory), let me know in the comments.
Part II, which covers intramuscular acidosis, can be found here.













I found this series very helpful! I appreciated the underlying chemistry explained in the first article in the series.
Thanks, and I’m glad it helped! I think it’s pretty interesting to look at some of these physiology questions in terms of biochemistry—it reveals a lot about what works (or doesn’t) and the root physical reasons for why.
“but never does the total size of that energy pool change.”
can you elaborate on that?
also i found it is curious that chris froome won tour de france 3 times with low carb diet
it is kind of hard to believe that he can do that with a disadvantage
What I mean is that diet cannot change the total size of our aerobic energy pool—only training can. In other words, if your total aerobic energy pool is capable of producing 750 kcal of work per hour, reducing carbohydrates will not increase or decrease your aerobic ability, it’ll only change how many of those 750 kcals of work are powered by carbohydrates (fewer) or fat (more). I write this mainly to combat the common notion that becoming “fat-adapted” means you’ll lose body fat easier or that you will increase your ability to do work within the aerobic zone, because neither are true. If you burn more body fat during exercise but don’t change total calories in or out, all that tells us is that you’re also storing more fat after meals. There’s nothing wrong with that, of course—our body stores fat to pull back out and use—but it’s important not to get sucked into the “fat-adapted means less fat on the body” mentality, because there’s no evidence that’s true or even a plausible mechanism for why it might be.
As for Chris Froome (now the 4-time winner of the Tour de France), there is a lot of misinterpretation based on a quick interview in which he basically says he lost weight in the off-season with a lower-carb (but not necessarily low-carb) diet. Reading through the various interviews he’s given, and also looking at what the Team Sky sports nutritionist says his athletes eat during the Tour itself, it’s clear that Froome still eats considerably more carbs than the typical athlete, he’s just a bit more picky about them now. Specifically, on “intense” days he’s cited to consume between 6 and 10 grams of carbohydrate per kilo of bodyweight, which is right at the upper-end of most recommendations for carb consumption. I can’t find any information on what his intake would be on low or moderate days (the nutritionist says he tries to get the riders to burn fat and protein during lower intensity stages), but most likely we’re still looking at the 4-7 g/kg bodyweight range because the riders (Froome included) still need to maintain glycogen even if they need fewer exogenous carbs to power their riding. At any rate, the bottom line is that Froome eats an extremely high-carb diet during the Tour de France, exactly like every other rider at that level.
(Also, in general anecdotes aren’t very informative because athletes could be unaware of the true composition of their diet, could be deceitful about it for purposes of image, or could simply be a genetic anomaly. I have no doubt there are a small number of humans who perform as well or better on a ketogenic diet, but I also don’t doubt that 99%+ of people who try will get weaker for it and so I don’t recommend it en masse.)
thanks for the detailed explanation! it makes sense
the team sky article mentioned use protein as energy source which seems not covered in this series
does it mean the same thing as use fat as aerobic energy source?
also as the article suggested, riding on fat/protein yields better muscle adaptation
do you have any idea why?
another question is why they do that also not only in training but also racing?
is there other benefits?
I didn’t include protein because our body doesn’t use amino acids directly to generate energy, but rather converts them to either glucose (glucogenic amino acids) or ketone bodies (ketogenic amino acids) first and then metabolizes those molecules for energy. The conversion isn’t free, however, so burning protein is actually very inefficient for athletes (though for weight loss, that inefficiency is helpful). When Team Sky says they use protein as fuel, I would argue that they’re actually misconstruing what is really being burned—the protein may be a minor part, but what is really happening on less intense stages is a shift from majority carbohydrates (as necessitated by the time spent doing anaerobic work) to more fat oxidation (though probably still mostly carbs) and a bit of spare amino acid oxidation. More importantly, by keeping protein high, they prevent dips in plasma amino acid levels would could lead to central fatigue, so even if they’re not burning the amino acids they are contributing to the athletes’ endurance.
As for adaptation, it’s a bit harder to answer because it seems like the original intent may have been lost in the writing of the article (presumably by someone who does not have a strong background in sports science). If I were to make an educated guess as to what the article meant, it would be this: there is a limited amount of research suggesting that occasionally exercising in a glycogen-depleted state can cause an increase in fat-burning adaptations. For endurance athletes, this is key as their goal is to be as efficient at burning fat while simultaneously maintaining their ability to burn carbs well. If you train in a glycogen-depleted state (or on a low-carb diet) too frequently, you start to disfavor carbs as fuel and can not use them as well. Conversely, if you only ever train on muscles fully-stocked with glycogen and with a sports drink for exogenous carbs, you risk downplaying your ability to burn fat. By balancing the two, athletes are able to balance their body’s substrate of choice, improving performance long-term. On a race day, though, there’s no question that carbs will bring out the best performance—you just want to be sure that your training hasn’t mitigated your ability to burn fat, as well!
All of this is perhaps more important to endurance athletes than climbers, however, as climbers are much more reliant overall on anaerobic energy and use aerobic energy production mostly to replenish those anaerobic sources. It may be possible that training occasionally in a glycogen-depleted state would improve power-endurance, but it could also be so minimal in an overall anaerobic sport as to not be beneficial at all. Certainly, I would be eager to see such a trial done—and really, this points to why it’d be nice to see more climbing-specific research in general!
Hi,
I wanted to address your statement:
“Thus, it’s difficult to imagine a way in which ketosis would improve performance”
I guess the main argument here would be that we can run out of glycogen but it takes a lot longer until we run out of fat. I don’t mean to disagree with any of your scientific arguments and conclusions nor am I recommending a ketogenic diet but I can see why some would trade in anaerobic glycolysis for not having to worry about bonking. Your argument seems to rely on readily available carbs which may not be realistic in all circumstances. And I’m not only talking about long trips through the mountains or physical activity that doesn’t allow sufficiently high carb intake such as ultra trail running – I sometimes find it difficult to get my fuel timing right for exercise during a working day.
Does that make sense?
Readily available carbs aren’t a prerequisite for preventing a bonk, they just ensure that bonking rarely or never happens. Nearly everyone, trained or not, will have enough glycogen stored to exercise for at least 2-3 hours without developing hypoglycemia. Athletes who must go more much longer than 3 hours generally train to improve not only their physical fitness, but also their glycogen utilization—they become better at making glycogen stretch further by ramping up fat burning. Going on a long trip through the mountains or an ultra trail run without training for it would be as inadvisable on a ketogenic diet as it would be on a normal one, threat of bonk or no!
Ketogenic diets essentially force a ramp-up of fat metabolism that could normally be achieved through training and remove the advantage of having those carbohydrates available in general. It may work for some people in some circumstances, but if performance is the goal, then you’re making a very real trade-off to avoid a situation [bonking] that is easily avoidable through proper preparation (including foreplanning about the foods you’ll bring on any extended endeavor).
And by ‘ramp up fat burning’ you refer to increasing aerobic metabolism in general and training in a fasted state?
I mean shifting our aerobic metabolism more towards the “burning fat” part of the spectrum rather than the “burning carbs” part. As we train (aerobically), we become both more efficient and able to burn more calories at once (through increased mitochondrial density, for example); as this happens, the overall aerobic benefits of carbohydrates (improved oxygen efficiency, improved ATP regeneration speed) become less important, at least when within the aerobic window. As a result, our body will start burning fat to a greater degree and reserving carbs to the extent it can. With ketosis, we essentially force that transition—our body becomes better at burning fat, but it also “forgets” how to burn carbs and so we don’t get the same type of benefit.
Thanks a Lot. Straightforward explanation of energy metabolism but there are some pieces of info i still need to fix.
“but never does the total size of that energy pool change. We merely train our body to burn one fuel better while it simultaneously “forgets” how to burn the other fuel”
Could you please give us any reference.
Acording to that ketogenic diets have never demonstrated to improve performance I would give you one
Keto-adaptation enhances exercise performance and body composition responses to training in endurance athletes
Fionn T. McSwiney, Bruce Wardrop, Parker N. Hyde, Richard A. Lafountain, Jeff S. Volek, Lorna Doyle
doi: 10.1016/j.metabol.2017.10.010
Could you explain us the results?
In regards to your first point, there are two ideas at play:
1. Fuel availability determines fuel utilization. On a high-carb diet, our body burns more carbs; on a high-fat diet, our body burns more fat. Essentially, our body burns whatever fuel we provide it in whatever proportion we provide it in, and it does this without significantly altering our total daily energy expenditure. This is critical, since if changing our diet changed how much energy our body needed to survive, there would be numerous hard-to-solve energy balance issues as some diets could hypothetically provide a “normal” amount of energy but also reduce our need for energy, making it so we gained weight rapidly (or vice-versa). Thus, conservation of energy expenditure is built-in to our physiology. Here’s a reference; there are many more that demonstrate something similar, but at this point it’s textbook knowledge.
2. Specifically for exercise, most studies on fat adaptation that have also examined subsequent carb refeeding have noticed that carbs are “spared” even after they return to the diet. Initially this was thought to be muscle glycogen sparing, but further research revealed that it was more akin to muscle glycogen impairing—our muscles downregulated their ability to burn carbs as they upregulated their ability to burn muscle. This is the same phenomenon as pointed out in the above point, and the bottom line is we can’t trick our body into burning more fuel. The best we’ve seen is that by providing exogenous carbs within an already high-carb diet can provide a very minimal boost to performance, but only in long-distance endurance events. Otherwise, our body uses the energy it has available, again in proportion to its availability. If we make fat very available and carbs not available, we burn fat readily but not carbs. If we change our diet, our body adapts, but never does it reach a point where we’re burning more energy than normal. Here’s a good review article covering some of the history of this concept in sport.
In regards to your second point, the study you reference didn’t demonstrate improved performance, it only showed that there was no loss in performance. This is a common finding, but often shadowed by loss in sprint performance. This study attempted to demonstrate that sprint performance wasn’t impaired (and even improved), but they used a laughably short 6 seconds as their time. How many cyclists need to sprint for only 6 seconds in a 100 kilometer race? As the study noted, 6 seconds is short enough that all the energy used will come from the creatine phosphate system, so no impairment would be expected, but overall nothing really changed either—while the ketogenic group had a higher peak power, their average power was statistically the same as the carb group, and it’s average power that makes a bigger impact in a sprint (as compared to, for example, a max lift).
The ketogenic group did lose more weight than the carb group, but that’s probably because the average body fat percentage of the carb group was 12.8% at the beginning while the ketogenic group was 17.5%; both groups ended up at about the same percentage (12.1 vs. 12.3%). The authors claimed this demonstrates that the ketogenic diet is a good way for some individuals to maintain race weight, and I don’t disagree—but they didn’t show that a normal- or high-carb diet doesn’t do the same since the carb group started at a normal bodyweight and didn’t need to lose weight.
Certainly, the ketogenic diet is going to work (or appear to work) for some people. For others, it might help them lose weight, and that could improve performance. When looked at in aggregate, though, there’s no evidence that it improves performance, only that it can maintain it, and only then if you ignore a loss in power in the 10 second to 2 minute range (anaerobic glycolysis). So, I hesitate to recommend ketogenic diets, because massively changing one’s diet without seeing significant benefits in terms of performance isn’t worth it.
Hope that helps, thanks for the comment!